 Rehabilitáció csak online elérhető
Rehabilitáció csak online elérhető
 E-mail: paleomedicina@gmail.com
E-mail: paleomedicina@gmail.com


A tanulmány áttekinti azokat az érveket, amelyek szerint az alvás szakaszai sajátos mechanizmust képeznek a tanulási és emlékezeti konszolidáció számára. A REM-alvás szerepéről számos ellentmondó elképzelés született. Ugyanakkor az alvási orsóknak topografikusan is sajátos szerepét sikerült megnyugtatóan tisztázni a konszolidációban. A bal oldali frontális orsók például a szerzők saját adatai szerint is a verbális megőrzésben játszanak központi szerepet.

CLEMENS ZSÓFIA – HALÁSZ PÉTER
Országos Pszichiátriai és Neurológiai Intézet
Magyar Pszichológiai Szemle, 2008, 64. 1. 19–28. DOI: 10.1556/MPSzle.64.2008.1.3.
Szinte az állatvilág egészére jellemző, hogy tagjai időről-időre megszakítják a külvilággal való kapcsolatukat, valamilyen nyugalmi állapotba merülnek, melynek ideje alatt testmozgásaik felfüggesztődnek, környezetük iránt való receptivitásuk lecsökken, miközben agyukban komplex idegrendszeri folyamatok zajlanak. Az alvás általános és evolúciós szempontból konzervatív jelenség, ami azt jelenti, hogy más viselkedésformákkal összehasonlítva az evolúció során keveset változott (BÓDIZS, 2000). Úgy tűnik, hosszabb ideig nélkülözhetetlen is, mivel például két hétig tartó teljes alvásmegvonás már a kísérleti állatok halálát okozhatja. Számos közvetlen és közvetett módon nyereségként elkönyvelhető funkciót lehet az alváshoz rendelni, mint például az energiamegtakarítást, az energialeadás csökkentését, a potenciális éjszakai ragadozók és más veszélyforrások elkerülését, a napközben elhasználódó fizikai-kémiai folyamatok regenerációját és az információfeldolgozás különféle formáit, például a memóriakonszolidációt. Könnyen belátható, hogy a felsorolt funkciókkal kapcsolatos nyereségek egy része alvás nélkül, pusztán pihenéssel is elérhető lenne. A memóriakonszolidáció azonban, mint a későbbiekben látni fogjuk, olyan funkció, amely magyarázhatja az alvás alatti tudatvesztés szükségességét és az alvó agyban zajló idegélettani folyamatok komplexitását.
Memóriakonszolidáció alatt kétféle jelenséget értünk. Az egyik arra vonatkozik, hogy a tanulást követő alvás során az emléknyomok megerősödnek, aminek eredményeképpen azután könnyebben hozzáférhetővé válnak és az interferáló ingereknek is jobban ellenállnak. A másik, hogy az emléknyomok neuronális reprezentációi fizikailag is átszerveződnek és az átmeneti memóriatárhelyekről a stabilabb és tartós megőrzésre alkalmas tárhelyekre kerülnek. Információelméleti megfontolások alapján arra következtettek, hogy mivel a külvilág ingereinek online feldolgozása, beleértve a tanulást is, és az emléknyomok ugyanazt a neuronális hálózatot használják, a rendszer limitált kapacitása mellett, egyidejűleg nem funkcionálhatnak hatékonyan. Eszerint az alvás- és ébrenlétszakaszok szabályszerű váltakozása tulajdonképpen olyan mechanizmus, amely lehetővé teszi, hogy a tanulás és konszolidáció folyamatai időben elkülönüljenek és egymással ne interferáljanak (MCCLELLAND, MCNAUGHTON, O’REILLY, 1995).
Az alvás és memória közti kapcsolat tudományos kutatásának története Jenkins és Dallenbach 1924-ben elvégzett úttörő kísérletéig nyúlik vissza (JENKINS, DALLENBACH, 1924). Kísérleti alanyaiknak értelmetlen szótagokat kellett megtanulniuk, melyeket 2, 4, 6, illetve 8 órás késleltetést követően kellett felidézniük, mely időszakot ébren vagy alvással töltötték. Kimutatták, hogy hasonló hosszúságú intervallumok mellett a tanultak felidézése sokkal hatékonyabb volt, ha a kísérleti személyek átaludták a szóban forgó intervallumokat, mint ha ugyanazt az időszakot ébren töltötték. Ezek a kísérletezők még nem tulajdonítottak aktív szerepet az alvásnak. Eredményeiket azzal magyarázták, hogy mivel az alvás egy időre kizárja az új élményszerzést, így interferáló ingerek hiányában az alvást megelőző emlékek is jobban megőrződnek.
REM-ALVÁS ÉS EMLÉKEZETI KONSZOLIDÁCIÓ
Az alváskutatás legnagyobb jelentőségű felfedezésének tartják a REM-alvás, azaz a gyors szemmozgásokkal és álomélményekkel jellemezhető alvásfázis 1953-ban történt felfedezését. Ekkor ismerték fel azt is, hogy az alvás tulajdonképp két, egymástól élettani szempontból gyökeresen eltérő állapot, a REM-alvás és a lassú hullámú alvás (LHA) váltakozásából épül föl. Ez a felfedezés az alvás és memória kapcsolatának kutatásában is fordulópontot jelentett. Ekkor jelentek meg az első elméletek, melyek a REM-alvásnak aktív szerepet tulajdonítottak az emléknyomok megőrzésében. Ezek az elméletek a freudi álomelméletből is merítettek, noha Freud maga sosem állította, hogy az álmok a memóriakonszolidáció szolgálatában állnának (VERTES, 2004).
Az emlékezeti konszolidáció REM-függésének vizsgálata a hatvanas, hetvenes évek alváskísérleteinek fő gerincét adta, és kezdetben ígéretes vonalnak tűnt. A memóriakonszolidáció REM-függésének elméletét megalapozó fő bizonyítéknak tekintették, hogy patkányokban a sikeres tanulást követően REM-alvásfokozódást figyeltek meg, valamint hogy a tanulást követő alvásban a REM-alvás megvonása Az alvás és az alvás alatti emlékezeti konszolidáció a memóriateljesítmény egyértelmű romlását vonta maga után. Ezt az összefüggést a tanulás számos típusában, így például az elkerülő tanulás, az operáns kondicionálás és a helytanulás különböző eseteiben is érvényesnek találták (SMITH, 1995). REM-alvás mellett szóló bizonyítékként könyvelték el azokat az eredményeket, miszerint ebben az alvásfázisban kondicionált válaszokat lehet kiépíteni sejtszinten, szemben a lassú hullámú alvással, ahol ez a kapcsolat nem alakul ki; továbbá, hogy az ébrenlét során kiépített kondicionált választ REM-alvásban is ki lehet váltani (MAHO, BLOCH, 1992; HENNEVIN, MAHO, HARS, 1998).
További támogatást jelentettek a REM alatti ún. „cueing” módszer alkalmazása során nyert eredmények. Ennek lényege, hogy ha a tanulást követő REM-alvásban kondicionáló ingerrel stimulálják az állatot, ez a korábban tanult ingerkapcsolat megerősödését eredményezi (HARS, HENNEVIN, PASQUES, 1985). Támogató adatként értelmezték, hogy REM-alvásban a tanulásra jellemző 4–8 Hz-es théta frekvenciájú aktivitás újra jelentkezik a hippocampusban. A hippocampális théta ritmusról ismert, hogy a tanulásban részt vevő idegsejtek aktivitását időzíti, és evvel a tanulás idegélettani alapjának tekintett hosszú távú potenciáció szolgálatában áll. Ennek alapján feltételezték, hogy a hippocampális thétának a REM-alvás alatti visszatérése a tanult emléknyomok egyfajta reaktivációját tükrözi.
Sajátos fordulat, hogy a memóriakonszolidációnak a REM-alvással kapcsolatos elmélete utólag zsákutcának bizonyult. Kiderült ugyanis, hogy a tanulási kísérletekben a REM-mel való összefüggés elsősorban a tanulási helyzetek és a REM megvonás külön-külön is erősen stresszkeltő hatásának volt köszönhető. Ezt támasztja alá, hogy a REM szerepe szempontjából pozitívnak bizonyult tanulási helyzetek túlnyomó többsége az averzív tanulási típusok körébe sorolható, és csak kevesebb köztük az apetitív tanulási forma. Állatkísérletekben a REM-alvás szelektív megvonásának céljából alkalmazott állványtechnika lényege, hogy a kísérleti állatok egy vízbe helyezett szűkös állványon mindig csak a REM-alvásuk beálltáig aludhattak, amikor is izomtónusuk megszűnése miatt lecsúsztak az állványról, vízbe estek és fölébredtek. Más kérdés, hogy a REM-megvonás módszertől függetlenül, tehát a farmakológiai REM-megvonás esetében is, stresszkeltő hatású lehet.
Komoly interpretációs nehézséget okozott az is, hogy a REM-mel kapcsolatos pozitív eredményeket emberben csak olyan ritka esetekben sikerült megismételni, mint például egyetemisták év végi vizsgaidőszaka, intenzív idegennyelv-tanulás, vizuális inverziót okozó szemüveg viselése és egy komplex logikai feladat (a Hanoi torony) tanulása. A REM-depriváció viszont következetesen nem okozott memóriakonszolidációs veszteséget a verbális és a téri-vizuális tanuláspróbákban (RAUCHS, DESGRANGES és munkatársai, 2005). Látványos adalékot jelentett azoknak a depressziós betegeknek az esete, akiknél az antidepresszáns kezelés hónapokra vagy akár egy teljes évre is kiterjedő teljes REM-hiányt idézett elő, de a betegek ennek a kognitív teljesítőképességükre gyakorolt észrevehető negatív hatását nem észlelték (VERTES, EASTMAN, 2000).
A REM-alvással kapcsolatos ellenmondásokat később az emberi memóriafolyamatok implicit/explicit kettősségére vonatkozó koncepció alkalmazásával próbálták föloldani. Ennek alapjául elsősorban Jan Born munkacsoportja által az 1990-es években végzett kísérletsorozat eredményei szolgáltak (PLIHAL, BORN, 1997, 1999). Ezekben a kísérletekben az éjszaka első mélyalvásban és a második REM-alvásban gazdag felének az emlékezésre gyakorolt hatását hasonlították össze. Kimutatták, hogy ha a kísérleti személyek alvását az éjszaka első felére korlátozzák és a második felében virrasztaniuk kell, ez nem érinti a verbális és vizuális deklaratív memóriájuk konszolidációját, viszont károsodik az implicit tanulás. Fordított esetben viszont elsősorban a deklaratív memória károsodik, de az implicit konszolidáció nem. Ennek alapján feltételezték, hogy a REM és a LHA különböző mértékben járulnak hozzá a hippocampustól függő (deklaratív: azaz az eseményekkel és tényekkel kapcsolatos) és a hippocampustól független, non-deklaratív (implicit: azaz a gyakorlással elsajátítható és tudatosan hozzá nem férhető) emléknyomok, készségek megőrzéséhez. Míg az implicit memóriaformák a REMalvásból profitálnak, a deklaratív emlékezést a lassú hullámú alvással hozták összefüggésbe.
Az újabb eredmények figyelembevételével és a REM-deprivációval kapcsolatban megfogalmazott módszertani kritikák figyelembevételével azonban úgy tűnik, még tovább kell szűkíteni azoknak a tanulási formáknak a körét, melyek valóban REM-függést mutatnak. Az ismételt vizsgálatok kimutatták, hogy több, korábban implicit jelenségkörbe sorolt tanulási típus, mint például a vizuális diszkriminációs készség (GAIS, PLIHAL és munkatársai, 2000) és motoros procedurális készségek konszolidációja inkább LHA-függést mutat, más implicit feladattípusok pedig mind a REM-től, mind a LHA-tól vagy akár az alvás egészétől is függetlennek bizonyultak (VERTES, 2004).
Talán csak egyetlen humán tanulási forma maradt, ami beilleszthető a REMfüggő tanulás koncepcióba, ez pedig a negatív érzelmi tónussal társuló emléknyomoknak a konszolidációja. Egy kísérletben egy film erősen stresszkeltő jeleneteihez való adaptációt vizsgálták. Kimutatták, hogy amennyiben az első bemutatást követő éjszaka során REM-megvonást alkalmaztak, a film újbóli megtekintése során a vártnál kevésbé csökkentek a stresszkeltő jeleneteket kísérő vegetatív reakciók, vagyis károsodott a stresszhez való adaptáció (GREENBERG, PILLARD, PEARLMAN, 1972). Egy másik kísérletben azt igazolták, hogy az éjszaka második, REM-alvásban gazdagabb fele a negatív emocionális tónusú szöveg megjegyzésének kedvez (WAGNER, GAIS, BORN, 2001). Egy harmadik kísérletben énfenyegető érzelmi terhelést jelentő anyag megjegyzése bizonyult REM-alvás megvonására érzékenynek (GRIESER, GREENBERG, HARRISON, 1972).
Feltételezhető, hogy a REM-alvás szerepe egy általánosabb érzelmi feldolgozásban van, aminek részjelensége a negatív érzelmi tónusú emléknyomok tanulásának REM-alvással való összefüggése. Erre utal, hogy a REM-alvás megvonása egészséges személyeknél hangulatromboló hatású, valamint hogy a REM-alvás fokozódását, cirkadián és egyéb eltéréseit depresszióban és poszttraumás stresszbetegségen is leírták. Azt is feltételezik, hogy a poszttraumás stresszbetegségben a kényszerűen visszatérő kínzó emlékezést és a traumával kapcsolatos rémálmok állandósulását ezeknek az emlékeknek a REM-mel kapcsolatos „túlkonszolidációja” okozza. Mindenesetre az érzelmi inputok feldolgozásában kulcsszerepet játszó amigdalának a REM-alvás idején tapasztalható fokozott aktivitásával ezek az adatok összhangban vannak (MAQUET, PÉTERS és munkatársai, 1996).
A LASSÚ HULLÁMÚ ALVÁS ÉS AZ EMLÉKEZETI KONSZOLIDÁCIÓ
Az újabb adatok fényében az implicit memória REM-függésének koncepciója tehát gyengülni látszik, a deklaratív memória LHA-sal való kapcsolatáról szóló feltételezés viszont folyamatosan erősödik. Ráadásul egyre több implicitnek tekintett memóriatípusról derül ki, hogy konszolidációja a LHA-tól függ. Egy implicit motoros adaptációs feladat esetében, például a feladat gyakorlását követő alvásban a lassú hullámú aktivitás fokozódását találták a jobb parietális lebeny egy jól körülírható régiójában, ráadásul ez az aktivitásfokozódás a készségnek az alvás alatti javulásával is összefüggést mutatott (HUBER, FELICE GHILARDI és munkatársai, 2004).
Egy másik kísérletben funkcionális képalkotó vizsgálattal azt mutatták ki, hogy azoknál a kísérleti személyeknél, akik hosszabb időn keresztül egy virtuális navigációs feladatot gyakoroltak, a tanulást követő alvás lassú hullámú fázisaiban fokozódott a hippocampális aktivitás. Ráadásul ez az aktivitásfokozódás egyenes arányban állt az emlékezeti megőrzés hatékonyságával (PEIGNEUX, LAUREYS és munkatársai, 2004). Más kísérletekben a LHA-ra jellemző humorális környezet manipulálásának hatását vizsgálták. Ismert, hogy a lassú hullámú alvásra fiziológiás körülmények között az alacsony stresszhormon-szint jellemző. Kimutatták, hogy amennyiben alvás alatt a kortizolszintet megemelik, a korábbi kísérletekben LHA-függőnek bizonyult emléknyomok megőrzésének hatékonyságán is rontani lehetett.
Hasonló eredményt értek el a lassú hullámú alvás acetylcholin-szintjének manipulációjával, melynek során kimutatták, hogy a lassú hullámú alvás alacsony acetylcholin-szintje kritikus tényező a deklaratív konszolidáció szempontjából. Egy következő említésre méltó kísérletben MARSHALL, HELGADÓTTIR és munkatársai (2006) a tanulást követő LHA-ban a kérgi lassú oszcillációt utánzó, 0,75 Hz frekvenciájú gyenge transzkraniális egyenáramú stimulálációt alkalmaztak, ami az ingerlések közti periódusok lassú hullámú és orsóaktivitását, valamint az alvást megelőzően tanult verbális ingeranyag megőrzését egyaránt fokozta. A théta frekvenciának megfelelő 5 Hz-es stimuláció ugyanakkor hatástalan maradt (MARSHALL, HELGADÓTTIR és munkatársai, 2006).
AZ ALVÁSI ORSÓK SZEREPE
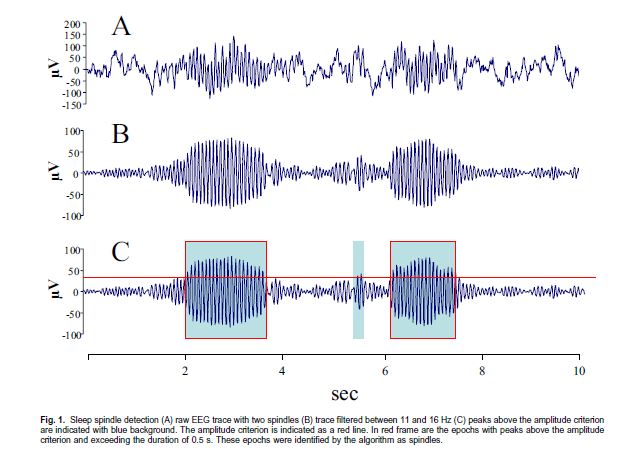
Alvási orsók, a kép forrása: Clemens et al., 2005. Overnight verbal memory retention correlates with the number of sleep spindles
Az alvási orsók a LHA jellegzetes, 11–16 Hz-es szinkron oszcillációi, melyek eredetüket tekintve a thalamocorticális rendszer oszcilláló üzemmódjának a termékei. Sejtszintű vizsgálatok kimutatták, hogy az alvásiorsó-aktivitás piramissejtekben a kálcium-ionok fokozott beáramlását idézi elő, ami további intracelluláris események láncolatát elindítva kedvez a hosszú távú plasztikus változásoknak (SEJNOWSKI, DESTEXHE, 2000). Felmerült, hogy a viselkedéses kísérletekben a LHA-ra jellemzőnek talált memóriakonszolidációs hatás vajon speciálisan köthető-e a LHA alatti alvásiorsótevékenységhez. Az alvási orsókat célzó vizsgálatok ezt támasztják alá: kimutatták például, hogy intenzív verbális és téri-vizuális tanulás az alvásiorsó-tevékenység fokozódását vonja maga után (MEIER-KOLL, BUSSMANN és munkatársai, 1999; GAIS, MÖLLE és munkatársai, 2002). Nemrég ezt egy motoros tanulás esetében (NISHIDA, WALKER, 2007), illetve állatokban (ESHENKO, MÖLLE és munkatársai, 2006) apetitíven kondicionált szagingerek tanulásával kapcsolatban is megerősítették. Hasonló következtetést lehet levonni azokból a kísérletekből, melyek a 2-es stádiumú alvásban töltött idővel találtak összefüggést a motoros tanulással kapcsolatban (WALKER, BRAKEFIELD és munkatársai, 2002).
Az alvási orsóknak a deklaratív memóriában betöltött szerepének bizonyításához saját humán vizsgálatainkkal is hozzájárultunk (CLEMENS, FABÓ, HALÁSZ, 2005; 2006). Olyan „szelíd” kísérleti elrendezést választottunk, melyben nem manipuláltuk az alvást, hanem csak az éjszaka folyamán előforduló alvási orsók számában meglévő individuális különbségek hatását vizsgáltuk. A vizsgálat tanulási fázisában arcokat és neveket mutattunk be az önkéntes kísérleti személyeknek. A nevekre való emlékezést szabad felidézés formájában kétszer ellenőriztük: először egy háromperces késleltetést követően, másodszor pedig az éjszakai alvást is magába foglaló 12 órás késleltetés után másnap reggel. Alvás alatti megőrzésnek definiáltuk a két szabad felidézés pontszámai közötti különbséget, amit az éjszakai alvás különböző változóival vetettük össze. Eredményeink szerint az alvás alatti verbális megőrzés a bal frontális területet lefedő elvezetésekben regisztrált alvási orsók teljes számával mutatott pozitív korrelációt. Az összefüggés nem volt szignifikáns sem a többi agyi területet lefedő elvezetésben regisztrált orsószámmal, sem az egyes alvásstádiumokban eltöltött idővel (1. ábra). Egy másik hasonló elrendezésű kísérletünkben egy téri-vizuális feladat, a Rey-Oesterrieth komplex ábra alvás alatti megőrzése a parietális területek fölött regisztrált összalvási orsószámmal mutatott összefüggést, de független volt a többi elvezetésben detektált orsószámtól és az egyes alvásstádiumokban töltött időktől is. Az 1. ábra az alvás alatti verbális megőrzés kapcsolatát mutatja az alvási orsók számával az egyes koponya felszíni elvezetési pontoknak megfelelően. Ugyanakkor az alvás alatti emlékezeti megőrzés a NREM-alvásban töltött teljes idővel mutatott szignifikáns összefüggést.
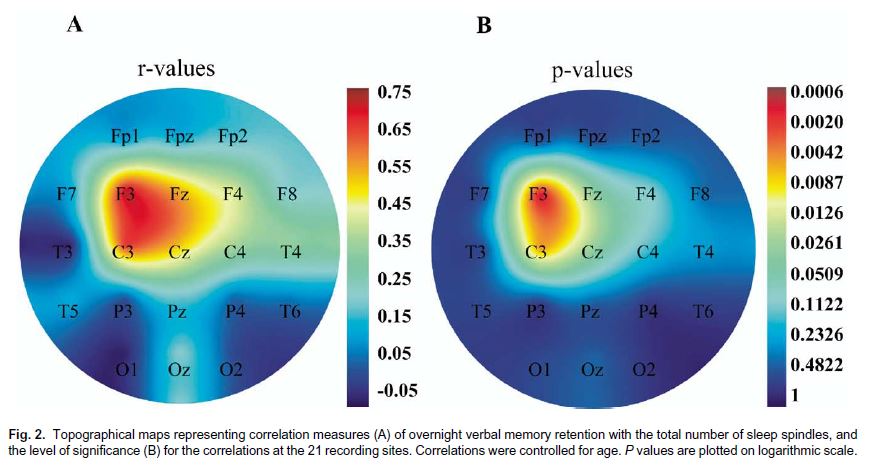
1. ábra. Az alvás alatti verbális emlékezeti megőrzés és az alvási orsók száma közti korrelációs koefficiensek topografikus térképe és a korrelációk szignifikanciája
Vagyis mind a verbális, mind a vizuális megőrzés kapcsolódik az alváshoz, de valószínűleg csak részben átfedő mechanizmusokhoz. Ezek az eredmények egyrészt megerősítették a lassú hullámú alvás szerepére vonatkozó elméleteket, ugyanakkor világossá tették, hogy a lassú hullámú alváson belül ez a pozitív hatás nem egyenletesen elosztott, hanem speciálisan az alvási orsókhoz köthető, mivel a LHA-stádiumokban töltött idővel kapcsolatosan ez az összefüggés nem volt szignifikáns. Ennek oka egyébként, hogy egyes emberek között az alvási orsók sűrűségében jelentős, akár többszörös különbségek is lehetnek. Másik fontos következtetés, hogy a frontális és parietális lokalizációjú alvási orsók eltérő mértékben vesznek részt a verbális és téri-vizuális modalitású ingeranyag konszolidációjában, amely ingeranyag specifikus topografikus megoszlás egyébként megfelel a bal frontális régiónak a verbális, illetve a perietális régiónak a téri-vizuális feldolgozásban betöltött ismert szerepének.
Az alvási orsók topografikusan specifikus szerepéhez további adalék, hogy a csak egyik kézzel végzett motoros tanulási feladat az ellenoldali félteke alvásiorsó-tevékenységét fokozta (NISHIDA, WALKER, 2007). A felsorolt eredmények, a lassú hullámú aktivitás hasonló szabályzására utaló (HUBER, FELICE GHILARDI és munkatársai, 2004) adatokkal együtt egy olyan lokális alvásteóriát körvonalaznak, melyben a szóban forgó aktivitások az adott funkció agyi lokalizációjának megfelelően homeosztatikus módon alkalmazkodnak a megnövekedett memóriakonszolidációs követelményekhez. A fenti tanulási komponenst tartalmazó példák esetében tapasztalatfüggő lokális homeosztázisról beszélhetünk. Fontos különbségtétel, hogy azokban az esetekben, amikor egy adott agyi területnek tanulástól független fokozott igénybevételről és ugyanennek a funkciónak az alvás alatti regenerációjáról van szó, használatfüggő lokális homeosztázisnak nevezik. Erre példa az egyik kéz (KATTLER, DIJK, BORBÉLY, 1994) vibrációs stimulálása révén kiváltott fokozott lassú hullámú aktivitás a szenzoros areáknak megfelelően.
A DEKLARATÍV EMLÉKEZETI KONSZOLIDÁCIÓ FUNKCIONÁLIS NEUROANATÓMIÁJA
A deklaratív emléknyomok alvás alatti konszolidációjával kapcsolatos magyarázatok központi eleme a hippocampus. A hippocampus filogenetikailag ősi struktúra, mely a kéreg valamennyi asszociációs és érző területével és a kéreg alatti struktúrákkal is kapcsolatban áll. BUZSÁKI György (1989) elmélete szerint az emléknyomok tartós rögzítése kétlépcsős folyamat eredménye. A tanulásnak megfelelő első szakaszban a szenzoros rendszerek felől érkező információk az entorhinális kéregben futó perforáns pályán keresztül a hippocampusba vetülnek, ahol egymással és a kéreg alatti inputokkal társítódnak. A hippocampus azonban csak átmenetileg tárolja ezeket az emléknyomokat: a második szakasznak megfelelő mélyebb konszolidáció alvásban történik. Az alvás alatti konszolidáció egy olyan off-line információfeldolgozási üzemmódnak tekinthető, melynek során az átmenetileg a hippocampusban tárolt reprezentációk átszerveződnek a stabilabb és tartósabb neocorticális tárhelyekre, a hippocampusból pedig törlődnek.
Ennek mechanizmusa a hippocampális-neocorticális dialógus, melynek lényege, hogy az ébrenlétre jellemző információáramlási irány LHA-ban a hippocampus afferens pályáinak gátlása és efferens pályáinak facilitációja révén megfordul, és a hippocampus felől a neocortex felé mutat. Kísérleti állatokban kimutatták, hogy a tanulásban aktív idegsejtek a LHA hippocampális meredek hullámainak idején újra aktiválódnak, és a tanulásra jellemző aktivitások térbeli-időbeli sorozatait gyorsított formában újra lejátsszák. Kimutatták továbbá, hogy a hippocampális meredek hullámok időben összerendeződnek a thalamocorticálisan generált alvási orsókkal (SIAPAS, WILSON, 1998; SIROTA, CSICSVÁRI és munkatársai, 2003). Feltételezik, hogy a hippocampális és neocorticális kódoló neuronok működése ezeknek a koordináltan előforduló oszcillációknak az útján hangolódik össze.
Fontos adalék, hogy míg korábban ilyen jellegű vizsgálatokat csak állatokban végeztek, nemrég epilepsziás betegek kivizsgálása során nyert intrakraniális regisztrátumokat felhasználva emberben is sikerült kimutatnunk a lassú hullámoknak, alvási orsóknak és a meredek hullámokkal együtt járó hippocampális fodroknak az időbeli összerendeződését (CLEMENS, FABÓ, HALÁSZ, 2007). Az epilepsziás betegek vizsgálata és gyógyítása számos olyan invazív beavatkozást tesz szükségessé, melyet – mintegy klinikai melléktermékként – alaptudományos kérdések vizsgálatára is fel lehet használni. Mivel a humán vizsgálatok lehetőségei nyilvánvaló etikai okokból korlátozottak, feltételezhető, hogy az epilepsziás betegek vizsgálata a jövőben is az alvás és memória kapcsolatára vonatkozó humán alaptudományos ismeretek egyik fő forrása lesz.
Mint ebből az összefoglalásból is kiderül, az alvás és memória közti kapcsolat kutatásának története nem egyenes vonalú történet. Jó példa erre a REM-alvással kapcsolatos érdeklődés felfutása, majd lecsengése vagy az alvási orsók szerepével kapcsolatos feltételezés, amely néhány évvel ezelőtt még értetlenséget váltott ki szakmai körökben, mára azonban az érdeklődés középpontjába került. Mivel a jelenségkörrel kapcsolatos kutatásokba egyre több kutatócsoport és diszciplína kapcsolódik be, további eredmények várhatók. Részleteitől függetlenül mára szinte dogmává vált az a vélemény, hogy az alvás alatt aktív emlékezetet konszolidáló folyamatok zajlanak. De mint általában, most sem árt vigyázni, amikor túl sokan kezdik ugyanazt gondolni. Az alvás alatti memóriakonszolidáció elméletének kisebbségben lévő kritikusai (SIEGEL, 2001) például cinikusan azt kérdezik, hogy egyáltalán minek tanulni, mikor ehelyett aludni is elég, utalván a memóriakonszolidációs elméletek népszerűségéből adódó nyilvánvaló elhajlásokra. Felvetik, hogy metodikailag kritizálható módon nyert eredmények is alapbizonyítékok közé keverednek, valamint, hogy az elmélet képviselői megfeledkeznek arról a hétköznapi, ugyanakkor nyilvánvaló tényről, hogy idővel az emléknyomok inkább halványulnak, mint erősödnek. Remélhető, hogy a jövőben a szofisztikáltabb vizsgálati módszerek előtérbe kerülésével pontosabban feltérképezhető lesz, hogy milyen korlátok között érvényesül az alvásnak a tanulásra és a felejtésre gyakorolt hatása.
SLEEP AND MEMORY CONSOLIDATION DURING SLEEP
CLEMENS, ZSÓFIA – HALÁSZ, PÉTER
The study surveys the arguments concerning the specific role of different sleep stages in memory and learning consolidation. There are several contradictory theories regarding the role of REM sleep. At the same time the topographically specific role of sleep spindles in consolidation was clarified. Left fontal spindles for example do play a central role in verbal retention according to data obtained by the authors.
Key words: sleep, learning, consolidation, sleep spindles, explicit and implicit memory
IRODALOM
BÓDIZS R.(2000) Alvás, álom, bioritmusok. Medicina, Budapest
BUZSÁKI, G. (1989) Two-stage model of memory trace formation: a role for “noisy” brain states. Neuroscience, 31, 551–570.
CLEMENS, Z., FABÓ, D., HALÁSZ, P. (2005) Overnight verbal memory retention correlates with the number of sleep spindles. Neuroscience, 132, 529–535.
CLEMENS, Z., FABÓ, D., HALÁSZ, P. (2006) Twenty-four hours retention of visuospatial memory correlates with the number of parietal sleep spindles. Neurosci Lett., 403, 52–56.
CLEMENS, Z., MÖLLE, M., EROSS, L., BARSI, P., HALÁSZ, P., BORN, J. (2007) Temporal coupling of parahippocampal ripples, sleep spindles and slow oscillations in humans. Brain, 130, 2868–2878.
ESCHENKO, O., MÖLLE, M., BORN, J., SARA, S. J. (2006) Elevated Sleep Spindle Density after Learning or after Retrieval in Rats. Journal Neuroscience, 26, 12 914–12 920.
GAIS, S., MÖLLE, M., HELMS, K., BORN, J. (2002) Learning-dependent increases in sleep spindle density. Journal Neuroscience, 22, 6830–6834.
GAIS, S., PLIHAL, W., WAGNER, U., BORN, J. (2000) Early sleep triggers memory for early visual discrimination skills. Nat. Neuroscience, 3, 1335–1339.
GREENBERG, R., PILLARD, R., PEARLMAN, C. (1972) The effect of dream (stage REM) deprivation on adaptation to stress. Psychosom Med., 34, 257–262.
GRIESER, C., GREENBERG, R., HARRISON, R. H. (1972) The adaptive function of sleep: the differential effects of sleep and dreaming on recall. Journal Abnormal Psychology, 80, 280–286.
KATTLER, H., DIJK, D. J., BORBÉLY, A. A. (1994) Effect of unilateral somatosensory stimulation prior to sleep on the sleep EEG in humans. Journal Sleep Res., 3, 159–164.
HARS, B., HENNEVIN, E., PASQUES, P. (1985) Improvement of learning by cueing during post-learning paradoxical sleep. Behaviour Brain Res., 18, 241–250.
HENNEVIN, E., MAHO, C., HARS, B. (1998) Neuronal plasticity induced by fear conditioning is expressed during paradoxical sleep: evidence from simultaneous recordings in the lateral amygdala and the medial geniculate in rats. Behaviour Neuroscience, 112, 839–862.
HUBER, R., FELICE GHILARDI, M., MASSIMINI, M., TONONI, G. (2004) Local sleep and learning. Nature, 430, 78–81.
JENKINS, J. B., DALLENBACH, K. M. (1924) Oblivescence during sleep and waking. American Journal of Psychology, 35, 605–612.
MAHO, C., BLOCH, V. (1992) Responses of hippocampal cells can be conditioned during paradoxical sleep. Brain Res., 581, 115–122.
MAQUET, P., PÉTERS, J., AERTS, J., DELFIORE, G., DEGUELDRE, C., LUXEN, A., FRANCK, G. (1996) Functional neuroanatomy of human rapid-eye-movement sleep and dreaming.
Nature, 383, 163–166.
MARSHALL, L., HELGADÓTTIR, H., MÖLLE, M., BORN, J. (2006) Boosting slow oscillations during sleep potentiates memory. Nature, 444, 610–613.
MCCLELLAND, J. L., MCNAUGHTON, B. L., O’REILLY, R. C. (1995) Why there are complementary learning systems in the hippocampus and neocortex: insights from the successes
and failures of connectionist models of learning and memory. Psychol. Review, 102, 419–457.
MEIER-KOLL, A., BUSSMANN, B., SCHMIDT, C., NEUSCHWANDER, D. (1999) Walking through a maze alters the architecture of sleep. Percept. Mot. Skills, 88, 1141–1159.
NISHIDA, M., WALKER, M. P. (2007) Daytime naps, motor memory consolidation and regionally specific sleep spindles. PLoS ONE 2, e341.
PEIGNEUX, P., LAUREYS, S., FUCHS, S., COLLETTE, F., PERRIN, F., REGGERS, J., PHILLIPS, C., DEGUELDRE, C., DEL FIORE, G., AERTS, J., LUXEN, A., MAQUET, A. (2004) Are spatial memories strengthened in the human hippocampus during slow wave sleep? Neuron, 44, 535–545.
PLIHAL, W., BORN, J. (1997) Effects of early and late nocturnal sleep on declarative and procedural memory. Journal Cognition Neuroscience, 9, 534–547.
PLIHAL, W., BORN, J. (1999) Effects of early and late nocturnal sleep on priming and spatial memory. Psychophysiology, 36, 571–582.
PLIHAL, W., BORN, J. (1999) Memory consolidation in human sleep depends on inhibition of glucocorticoid release. Neuroreport, 9; 10, 13, 2741-2747.
RAUCHS, G., DESGRANGES, B., FORET, J., EUSTACHE, F. (2005) The relationships between memory systems and sleep stages. Journal Sleep Res., 14, 123–140.
SEJNOWSKI, T. J., DESTEXHE, A. (2000) Why do we sleep? Brain Res., 886, 208–223.
SIAPAS, A. G., WILSON, M. A. (1998) Coordinated interactions between hippocampal ripples and cortical spindles during slow-wave sleep. Neuron, 21, 1123–1128.
SIEGEL, J. M. (2001) The REM sleep-memory consolidation hypothesis. Science, 294, 1058–1063.
SIROTA, A., CSICSVÁRI, J., BUHL, D., BUZSÁKI, G. (2003) Communication between neocortex and hippocampus during sleep in rodents. Proceeding Natl. Academic Science USA, 100, 2065–2069.
SMITH, C. (1995) Sleep states and memory processes. Behaviour Brain Res., 69, 137–145. VERTES, R. P. (2004) Memory consolidation in sleep; dream or reality. Neuron, 44, 135–148.
VERTES, R. P., EASTMAN, K. E. (2000) The case against memory consolidation in REM sleep. Behaviour Brain Science, 23, 867–876.
WAGNER, U., GAIS, S., BORN, J. (2001) Emotional memory formation is enhanced across sleep intervals with high amounts of rapid eye movement sleep. Learn Mem., 8, 112–119.
WALKER, M. P., BRAKEFIELD, T., MORGAN, A., HOBSON, J. A., STICKGOLD, R. (2002) Practice with sleep makes perfect: sleep-dependent motor skill learning. Neuron, 35, 205–211.
Kapcsolódó tudományos cikk
Baglyok és pacsirták, avagy mikor vegyünk pizsamát?
A Paleomedicina munkacsoport kizárólag tudományos alapon végzi a tevékenységét. Semmilyen természetgyógyászati módszert nem használunk, ezektől elhatárolódunk. A Paleomedicina által képviselt irányzat, az evolúciós orvoslás, a valódi tudomány része. Nemzetközi orvosi szakfolyóiratokban megjelent közleményeinket itt olvashatják.